C’était il y a 50 ans : la découverte de Lucy (5/6). La paléobiologie d’Australopithecus afarensis
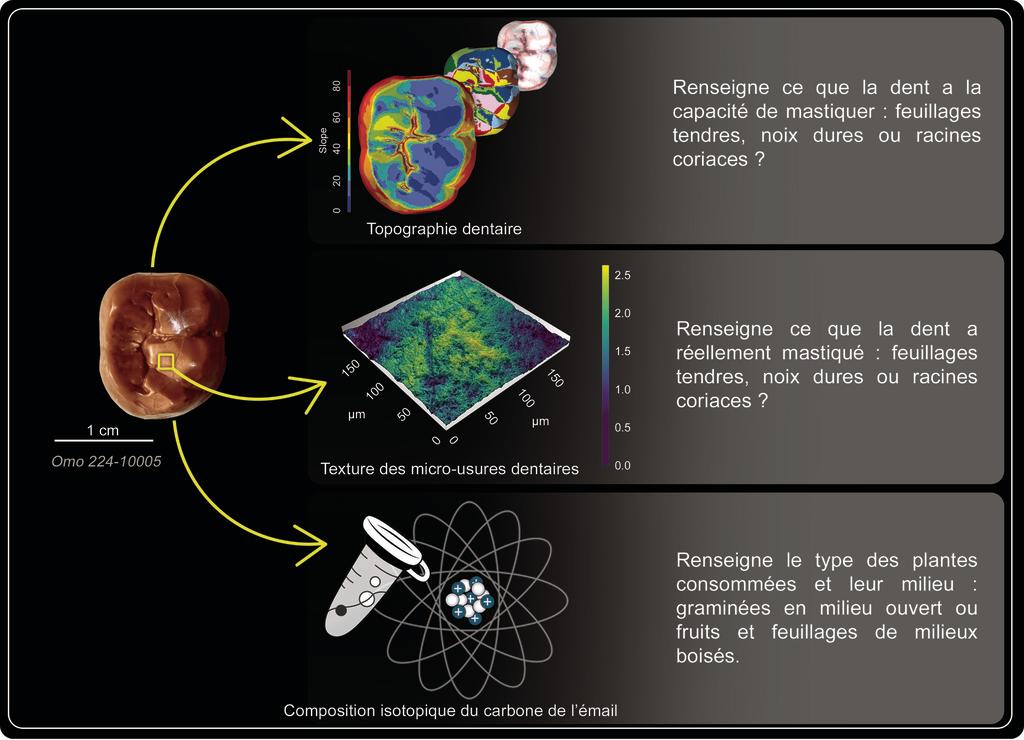
Méthode visant à caractériser les habitudes alimentaires d’A. afarensis. © Gildas Merceron, S. Riffaut, laboratoire PALEVOPRIM
Éthiopie, région de l’Afar, vallée de l’Awash : nous sommes en 1974, une équipe internationale vient de découvrir, à Hadar, le squelette fossile d’une espèce alors inconnue, Australopithecus afarensis. Ainsi est née Lucy, la plus célèbre de nos ancêtres. Pour fêter cet anniversaire sans nul autre pareil, Archéologia vous propose un vaste dossier présentant ces australopithèques qui ont vécu plus de 800 000 ans, entre 3,8 et 3 millions d’années (Ma). Connue aujourd’hui par de nombreux témoignages, cette espèce continue de nous livrer ses secrets. Venez découvrir qui étaient Lucy et les siens…
Les auteurs de ce dossier sont : Sandrine Prat, HNHP, CNRS, MNHN & UPVD, coordinatrice du dossier ; Jean-Renaud Boisserie, Centre français des études éthiopiennes, laboratoire PALEVOPRIM, CNRS, coordinateur du dossier ; Doris Barboni, Institut français de Pondichéry, CNRS & ministère de l’Europe et des Affaires étrangères ; Amélie Beaudet, laboratoire PALEVOPRIM, CNRS & université de Poitiers ; Gilles Berillon, HNHP, CNRS, MNHN & UPVD ; Raymonde Bonnefille, CNRS ; Marie Bridonneau, Centre français des études éthiopiennes, CNRS & ministère de l’Europe et des Affaires étrangères ; Franck Guy, laboratoire PALEVOPRIM, CNRS & université de Poitiers ; Donald Johanson, Institute of Human Origins, Arizona State University ; François Marchal, ADES, CNRS, Aix-Marseille université, & EFS ; Gildas Merceron, laboratoire PALEVOPRIM, CNRS & université de Poitiers ; Raphaël Pik, CRPG, CNRS & université de Lorraine ; Antoine Souron, PACEA, université de Bordeaux
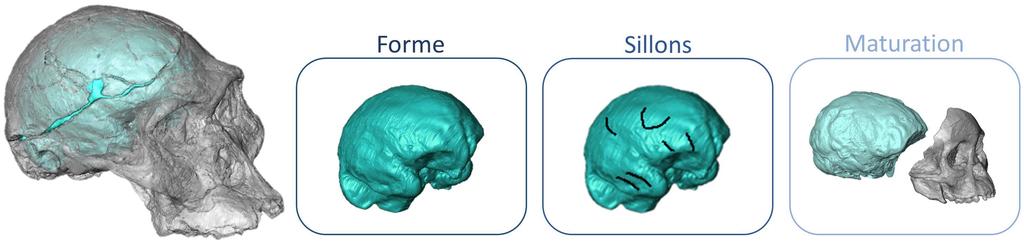
Niveaux de lecture des empreintes cérébrales préservées au sein d’un même fossile. © Amélie Beaudet
Notre capacité à nous déplacer et à interagir avec notre environnement physique et social est dépendante de l’énergie dont notre organisme dispose ; or celle-ci est puisée dans les ressources alimentaires. Que sait-on de ces dernières, consommées par Australopithecus afarensis et des impacts de ces comportements sur leur biologie ?
Le régime alimentaire d’A. afarensis
Comprendre l’écologie d’une espèce éteinte implique d’extraire des informations des tissus osseux et dentaires. Pour A. afarensis, elles sont principalement issues de ces derniers ; les dents sont en effet de véritables archives paléobiologiques, associées à une fenêtre temporelle donnée. Les molaires d’un australopithèque, combinant un émail plus épais et un relief dentaire moins élevé que les molaires des grands singes non humains actuels, suggèrent que les individus aux dents les plus résistantes aux fractures, associées à la consommation des aliments durs et coriaces (graines, racines, tubercules…), ont été sélectionnés au fil des générations. Si cette morphologie reste le premier indice sur la potentielle alimentation d’A. afarensis, elle renseigne sur un temps très long. L’usure des dents, et tout particulièrement la texture des micro-usures, complète nos connaissances. Ainsi la rugosité de la surface de l’émail des facettes masticatrices des molaires des australopithèques d’Afrique de l’Est est si faible que graines, racines, tubercules ne semblent, finalement, pas avoir été consommés, du moins au cours des jours ou semaines précédant la mort des spécimens étudiés (l’usure des dents étant continue, l’information extraite ne reflète qu’une période brève avant la mort).
Les analyses isotopiques éclairent l’écologie alimentaire d’Australopithecus afarensis.
L’émail en dit long
De son côté, l’émail renferme aussi des informations : ce tissu est en effet constitué d’éléments (carbone, oxygène, calcium, azote…) que l’organisme a inhalés en respirant ou ingérés en buvant ou absorbant les ressources à sa disposition. L’analyse de la composition isotopique de certains d’entre eux éclaire l’écologie alimentaire d’A. afarensis. Ainsi, la teneur des différents isotopes du carbone atteste la présence significative de plantes aujourd’hui principalement représentées par des herbacées des zones intertropicales, comme les graminées. A. afarensis sélectionnait ces herbacées mais aurait délaissé les parties dures comme les graines et les racines. Néanmoins, la morphologie des molaires confirme bien que ces hominines étaient en capacité de réduire des objets durs, moins accessibles, mais indispensables lors de saisons plus sévères.
Des méthodes similaires appliquées aux isotopes du calcium ont par ailleurs démontré que les éléments les plus nutritifs n’étaient pas toujours disponibles dans l’environnement ; ces fluctuations saisonnières se répercutaient par exemple sur la pratique de l’allaitement qui s’établissait sur une période plus ou moins longue. Les milieux boisés et clairsemés garantissaient l’accès aux ressources tout en fournissant l’énergie nécessaire aux activités quotidiennes et au maintien du bon fonctionnement de l’organisme. C’est particulièrement le cas du cerveau, organe énergivore qui joue un rôle central dans la perception et l’interaction avec l’environnement et a occupé une place déterminante dans l’évolution humaine.
Le cerveau d’Australopithecus afarensis
La comparaison du cerveau humain actuel avec celui des chimpanzés révèle une série de caractères propres au nôtre, comme une forme arrondie, une aire visuelle réduite, la présence de deux sillons dans la région de Broca ou encore une longue période de maturation après la naissance. Ces traits sont probablement à l’origine de comportements uniques, comme la fabrication d’outils ou le langage, en partie responsables de notre succès évolutif. Ces spécificités étaient-elles déjà présentes chez A. afarensis ? La réponse à cette question est loin d’être évidente en raison des problèmes pratiques qu’elle soulève. En l’absence de tissus mous, le cerveau des espèces fossiles est reconstruit à partir des empreintes qu’il laisse sur la surface interne de la boîte crânienne. En conséquence, seul l’aspect externe du cerveau, et donc sa forme ainsi que l’organisation des sillons cérébraux, est enregistré. Par ailleurs, nous disposons essentiellement de fragments d’os et de très peu de crânes complets d’A. afarensis, et plus largement d’Australopithecus. Néanmoins, en recoupant les informations disponibles pour l’ensemble de ce genre, les paléontologues ont établi que le cerveau de Lucy et des siens était relativement allongé, comme celui des chimpanzés actuels, et que l’aire de Broca ne montrait qu’un seul sillon.
Une maturation cérébrale plus longue
En revanche, le doute demeure sur l’architecture de l’aire visuelle, qui présentait une configuration soit humaine (réduite) soit proche de celle des chimpanzés (étendue). La découverte d’un crâne complet d’enfant d’A. afarensis a mis en évidence les premiers signes d’un allongement de la période de maturation du cerveau après la naissance. L’ensemble de ces changements a très certainement eu un impact sur l’énergie allouée au cerveau, probablement au détriment d’autres organes (comme ceux impliqués dans la digestion) et a donc modifié les comportements alimentaires sur le long terme. À l’heure actuelle, les implications fonctionnelles de ces descriptions sont encore incertaines. L’archéologie expérimentale et les neurosciences ont prouvé que les lobes pariétaux sont principalement sollicités dans les premières industries lithiques, mais les données fossiles sur cette région du cerveau sont presque inexistantes. Cependant, une période de maturation cérébrale plus longue a sûrement joué un rôle dans l’apprentissage de nouveaux comportements, notamment technologiques, et dans leur transmission.
Sommaire
C’était il y a 50 ans : la découverte de Lucy








